
NCS - MORFOLOGIA - 06/04/23 -SP 2.3
- thikow

- 6 de abr. de 2023
- 15 min de leitura
Atualizado: 24 de mai. de 2023
SP 2.3 ATÉ QUE ENFIM
Fertilização:
Fase 1
Reação de capacitação do espermatozóide: leva 7 horas, por isso que os primeiros não conseguem fecundar
Processo de capacitação: Remoção de membranas espermáticas, glicoproteínas, lipídeos para o espermatozoide fique apto a fecundação.
Reação acrossômica, na coroa radiata: Enzimas são catalisadoras, que quebram as estruturas das ligações da coroa, para alcançar a zona pelúcida.
Fase 2
Na zona pelúcida: Existe o receptor ZP3 ( so o espermatozoide tem). O primeiro espermatozóide que entra, ocorre a despolarização da zona pelúcida, assim, a célula fica mais negativa (mudanca de pólos) e nesse momento a zona pelúcida torna-se mais rígida. Evitando a fertilização por mais de 01 espermatozoide.
Fase 3
Espermatozoide entra, finalizando a meiose 2, formando o corpúsculo polar que será descartado.
Zigoto é formado. Fusão dos pro núcleos fem e masculinos. Tornando uma célula diplóide.
24 horas depois, ocorre a clivagem.
Blastômero - massa cheia de células embrionárias.
Mórula - absorve o glicogênio. Até o 5 quinto dia, a zona pelúcida é desfeita., formando blastocisto
Blastocisto - se caracteriza pela cavidade interna, camada externa que forma trofoblasto e cavidade interna o embrioblasto.
Nidação: processo de implantação no endométrio
Blastocisto vira embrioblasto, gera o embrião por si só.
Trofoblasto se divide em Citotrofoblasto e Sinciotrofoblasto - no futuro vira placenta.
Embrioblasto se divide em Epiblasto e Hipoblasto - cavidade do saco vitelínico.
Hexoceloma - Membrana, forma o saco vitelineo.
A) Quais os resultados da fertilização quanto ao número de cromossomos?
B) Em que momento se deu a determinação do sexo biológico do embrião?
C) O que são clivagens?
RA) 46 cromossomos
RB) Fase 3, fertilização na fase 3. Junção o do pro núcleo fem e masc e do gene.
RC) Clivagens: primeiras divisões mitóticas do desenvolvimento embrionário.
Modelo anatômico:
-Zigoto (embrião de 1 célula);
-Blástula (embrião de 2, 4, 8 células)
-Mórula (embrião de 16 células)
-Blastocisto (embrião com cavidade)
-Blastocele (cavidade do embrião)
-Massa celular interna
-Massa celular externa
-Epiblasto
-Hipoblasto
-Citotrofoblasto
-Sinciciotrofoblasto.





Ciclo menstrual
Na puberdade, a mulher começa a ter ciclos mensais regulares. Esses ciclos sexuais são controlados pelo hipotálamo. O hormônio liberador de gonadotrofinas (GnRH; do inglês, gonadotropin-releasing hormone), produzido pelo hipotálamo, age nas células do lobo anterior da adeno-hipófise, que, por sua vez, secreta gonadotrofinas. Esses hormônios, o hormônio foliculoestimulante (FSH; do inglês, follicule-stimulating hormone) e o hormônio luteinizante (LH; do inglês, luteinizing hormone), estimulam as alterações cíclicas dos ovários.
No início de cada ciclo ovariano o FSH estimula o crescimento de 15 a 20 folículos no estágio primário (pré-antral). Ele não é necessário para para promover o desenvolvimento dos folículos primordiais até o estágio de folículo primário, mas sem ele os folículos primários morrem e se tornam atrésicos. Sob condições normais apenas um desses folículos alcança a maturidade plena, e apenas um oócito é liberado; os outros degeneram entrando em atresia.
Quando um folículo se torna atrésico, o oócito e as células foliculares circunjacentes degeneram e são substituídos por tecido conjuntivo, formando um corpo atrésico.
O FSH também estimula a maturação das células foliculares (granolosas) ao redor do oócito, porém a proliferação dessas células é mediada pelo fator de diferenciação de crescimento 9, um membro da família do transformador do crescimento β (TGF-β).
Foliculos são estimulados pelo FSH a crescerem, e pelo FSH e LH a maturar. A oocitação acontece quando as concentrações de LH aumentam até altos níveis. O LH também promove o desenvolvimento do corpo lúteo.
Todos os dias alguns folículos primordiais de um conjunto começam a se desenvolver em folículos em crescimento, e esse crescimento é independente de FSH. Então, conforme o ciclo progride, a secreção de FSH recruta os folículos em crescimento a começarem a se desenvolver em folículos vesiculares (antrais). Durante os últimos dias de maturação dos folículos vesiculares, os estrógenos produzidos pelas células foliculares e da teca estimulam o aumento da produção de LH pela hipófise
, e esse hormônio faz com que o folículo entre no estágio vesicular maduro (de De Graaf) para completar a meiose I e, aproximadamente 3 horas antes da oocitação, entre na meiose II, ficando parado na metáfase.


A cada ciclo ovariano, alguns folículos primários começam a crescer, mas, em geral, somente um alcança a maturidade plena e apenas um oócito é liberado na oocitação. Na oocitação, o oócito está em metáfase na segunda divisão meiótica e está cercado pela zona pelúcida e por algumas células granulosas. O movimento das fímbrias tubais carrega o oócito para a tuba uterina.
Corpo lúteo
Após a oocitação, as células granulosas que permanecem na parede do folículo roto junto com as células da teca interna são vascularizadas pelos vasos circunjacentes. Sob a influência do LH, essas células desenvolvem um pigmento amarelado e se tornam as células luteínicas, que formam o corpo lúteo e secretam estrógenos e progesterona. A progesterona, junto com alguns estrógenos, faz com que a mucosa uterina entre no estágio progestacional ou secretor, preparando-se para a implantação do embrião.
Corpo albicans
Se a fertilização não ocorrer, o corpo lúteo alcança o máximo de desenvolvimento aproximadamente 9 dias após a oocitação. Ele pode ser facilmente reconhecido como uma projeção amarelada na superfície do ovário. Subsequentemente, o corpo lúteo encolhe por causa da degeneração das células lúteas (luteólise) e forma uma massa de tecido cicatricial fibrótico, o corpo albicans. Simultaneamente, a produção de progesterona diminui, causando o sangramento menstrual. Se o oócito for fertilizado, a degeneração do corpo lúteo é evitada pela gonadotrofina coriônica humana, um hormônio secretado pelo sinciciotrofoblasto do embrião em desenvolvimento. O corpo lúteo continua a crescer e forma o corpo lúteo gravídico (corpus luteum graviditatis). Por volta do fim do terceiro mês, essa estrutura pode ter de um terço até metade do tamanho total do ovário. As células lúteas amareladas continuam a secretar progesterona até o fim do quarto mês; desse ponto em diante, elas regridem lentamente, conforme a secreção de progesterona pelo componente trofoblástico da placenta se torna adequada para a manutenção da gravidez. A remoção do corpo lúteo gravídico antes do quarto mês em geral leva ao aborto.
Fertilização
Antes que os espermatozoides possam fertilizar o oócito, têm de passar por:
1. Capacitação, período no qual uma capa de glicoproteínas e de proteínas plasmáticas seminais é removida da cabeça do espermatozoide
2. Reação acromossômica, durante a qual são liberadas substâncias semelhantes à acrosina e à tripsina, para penetrar a zona pelúcida.
Durante a fertilização, o espermatozoide precisa penetrar na:
1. Coroa radiada
Até 500 milhões de espermatozoides mas somente um deles fertiliza o oócito. Acredita-se que os outros espermatozoides ajudem o fertilizador a penetrar as barreiras que protegem o gameta feminino.
2. Zona pelúcida
Camada de glicoproteínas que cerca o oócito, facilita e mantém a ligação do espermatozoide e induz a reação acrossômica. A permeabilidade da zona pelúcida se altera quando a cabeça do espermatozoide contata a superfície do oócito, liberando enzimas lisossomais dos grânulos corticais que por sua vez alteram as propriedades da zona pelúcida, evitando a possibilidade de penetração de qualquer outro esperatozoide.
3. Membrana celular do oócito.
A adesão inicial do espermatozoide ao oócito é mediada parcialmente pela interação de integrinas do oócito com seus ligantes, desintegrinas, no espermatozoide. Após a adesão, as membranas plasmáticas do espermatozoide e do oócito se fundem
Tão logo o espermatozoide penetre no oócito:
1. O oócito termina sua segunda divisão meiótica e forma o pró-núcleo feminino
2. A zona pelúcida se torna impenetrável a outros espermatozoides
3. A cabeça do espermatozoide se separa da cauda, incha e forma o pró-núcleo masculino.
Após os pró-núcleos replicarem seus DNAs, os cromossomos paterno e materno se misturam, dividem-se longitudinalmente e passam por uma divisão mitótica, dando origem ao estágio de duas células. Os resultados da fertilização são:
1. Restauração do número diploide de cromossomos
2. Determinação do sexo cromossômico
3. Início da clivagem.
A infertilidade é um problema para 15 a 30% dos casais e pode ser superada usando a tecnologia de reprodução assistida (TRA). A fertilização in vitro (FIV) envolve fertilizar os óvulos em um meio de cultura e colocá-los no útero no estágio de oito células. Em alguns casos, os óvulos são fertilizados pela injeção intracitoplasmática dos espermatozoides (ICSI), por meio da qual um único espermatozoide é injetado em um citoplasma do óvulo. Essas técnicas in vitroestão associadas a maior risco de defeitos congênitos, nascimentos prematuros, baixo peso ao nascer e nascimento múltiplo. Aproximadamente 1 a 2% de todos os nascimentos nos EUA ocorrem por TRA.
A clivagem é uma série de divisões mitóticas que resultam no aumento da quantidade de células, os blastômeros, os quais se tornam menores a cada divisão. Após três divisões, eles sofrem compactação, para se tornarem uma bola celular altamente agrupada com camadas interna e externa. Os blastômeros compactados se dividem para formar uma mórula de 16 células. Conforme a mórula entra no útero no terceiro ou no quarto dia após a fertilização, começa a aparecer uma cavidade, e se forma o blastocisto. A massa celular interna, que se forma por volta do período da compactação e que se desenvolverá no embrião em si, aloja-se em um dos polos do blastocisto. A massa celular externa, que envolve as células internas e a cavidade blastocística, formará o trofoblasto.
O útero está na fase secretória no período da implantação, e o blastocisto se implanta no endométrio ao longo da parede anterior ou posterior. Se a fertilização não ocorrer, então começa a fase menstrual, e as camadas endometriais esponjosa e compacta são liberadas. A camada basal permanece para regenerar as outras durante o ciclo seguinte.
O ÚTERO NO MOMENTO DA IMPLANTAÇÃO
A parede uterina consiste em três camadas:
1. Endométrio ou mucosa que reveste a parede interna
2. Miométrio, camada espessa de músculo liso
3. Perimétrio, o peritônio que reveste a parede externa.
Da puberdade (11 a 13 anos de idade) até a menopausa (45 a 50 anos de idade), o endométrio sofre variações em um ciclo de aproximadamente 28 dias sob o controle hormonal dos ovários. Durante o ciclo menstrual, o endométrio uterino passa por três estágios:
1. Fase folicular ou proliferativa
2. Fase secretória ou progestacional
3. Fase menstrual
A fase proliferativa começa no fim da fase menstrual, está sob a influência do estrogênio e acompanha o crescimento dos folículos ovarianos. A fase secretória começa aproximadamente 2 ou 3 dias após a oocitação, em resposta à progesterona produzida pelo corpo lúteo. Se a fertilização não ocorrer, o sangramento do endométrio (camadas esponjosa e compacta) marca o início da fase menstrual. Se a fertilização ocorrer, o endométrio ajuda na implantação e contribui para a formação da placenta. Mais tarde, na gestação, a placenta assume o papel de produção hormonal, e o corpo lúteo degenera.

No início da segunda semana, o blastocisto está alojado parcialmente no estroma endometrial. O trofoblasto se diferencia em uma camada interna, que prolifera ativamente, o citotrofoblasto; e em uma camada externa, o sinciciotrofoblasto, que invade os tecidos maternos.

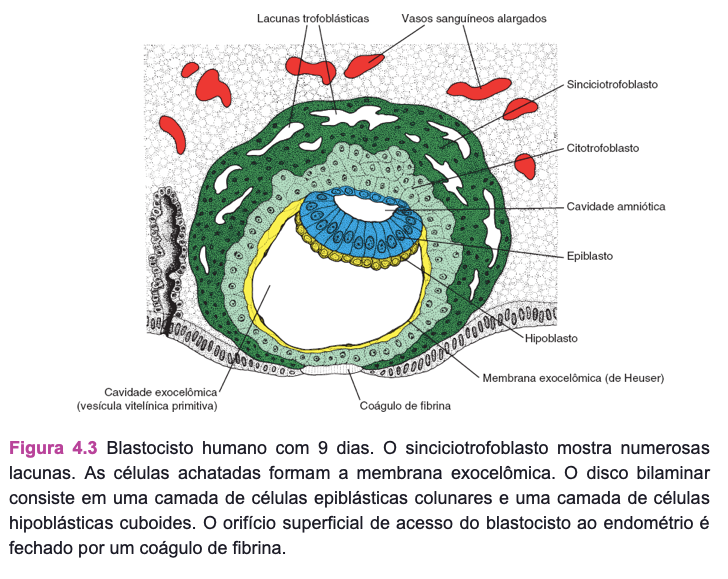


No nono dia, desenvolvem-se lacunas no sinciciotrofoblasto. Subsequentemente, os sinusoides maternos são erodidos pelo sinciciotrofoblasto, o sangue materno entra na rede lacunar e, por volta da segunda semana, começa uma circulação uteroplacentária primitiva. Enquanto isso, o citotrofoblasto forma colunas celulares que penetram e são cercadas pelo sincício, as vilosidades primárias. No fim da segunda semana, o blastocisto está completamente alojado e seu orifício de entrada na mucosa está fechado.


Enquanto isso, a massa celular interna ou embrioblasto se diferencia em epiblasto e hipoblasto, formando, juntos, um disco bilaminar. As células epiblásticas originam os amnioblastos que recobrem a cavidade amniótica, superior à camada epiblástica. As células hipoblásticas são contínuas com a membrana exocelômica, e, juntas, elas recobrem a vesícula vitelínica primitiva. No fim da segunda semana, o mesoderma extraembrionário preenche internamente o espaço entre o trofoblasto e o âmnio, bem como a membrana exocelômica. Quando se desenvolvem vacúolos nesse tecido, forma-se o celoma extraembrionário ou cavidade coriônica. O mesoderma extraembrionário que recobre o citotrofoblasto e o âmnio é o mesoderma extraembrionário somático; a cobertura que cerca a vesícula vitelínica é o mesoderma extraembrionário esplâncnico.
A segunda semana do desenvolvimento é conhecida como a “semana do dois”:
1.O trofoblasto se diferencia em duas camadas: o citotrofoblasto e o sinciciotrofoblasto
2.O embrioblasto forma duas camadas: o epiblasto e o hipoblasto
3.O mesoderma extraembrionário se divide em duas camadas: as camadas somática e esplâncnica
4.Formam-se duas cavidades: a amniótica e a da vesícula vitelínica.
A implantação ocorre no fim da primeira semana. As células trofoblásticas invadem o epitélio e o estroma do epitélio subjacente com a ajuda de enzimas proteolíticas. A implantação também pode ocorrer fora do útero, como na cavidade retouterina, no mesentério, na tuba uterina ou no ovário (gravidez ectópica).
O evento mais característico durante a terceira semana é a gastrulação. Ela começa com o surgimento da linha primitiva, que tem em sua parte cefálica o nó primitivo. Na região do nó e da linha, as células epiblásticas se movem para dentro (invaginam) para formar novas camadas, o endoderma e o mesoderma. As células que não migram através da linha primitiva, mas permanecem no epiblasto, formam o ectoderma. Assim, o epiblasto dá origem às três camadas germinativas, ectoderma, mesoderma e endoderma, e essas camadas formam todos os tecidos e órgãos.


As células pré-notocordais que invaginam na região da fosseta primitiva se deslocam para a região cefálica até alcançarem a placa precordal. Elas se intercalam no endoderma como a placa notocordal. Com a continuação do desenvolvimento, a placa se desprende do endoderma e forma um tubo sólido, a notocorda. Essa estrutura determina um eixo na linha média, que funcionará como a base do esqueleto axial.

As partes cefálica e caudal do embrião são estabelecidas antes que a linha primitiva tenha se formado. Assim, as células do hipoblasto (endoderma) na margem cefálica do disco formarão o EVA, que expressa genes de formação da cabeça, incluindo OTX2, LIM1 e HESX1, bem como o fator solúvel cerberus. NODAL, um membro da família de genes TGF-β é, então, ativado e inicia e mantém a integridade do nó e da linha primitivos. Na presença de FGF, o BMP4 ventraliza o mesoderma durante a gastrulação de modo que ele forme o mesoderma intermediário e o mesoderma da placa lateral. Chordin, noggin e follistatin antagonizam a atividade de BMP4 e dorsalizam o mesoderma para originar a notocorda e os somitômeros na região da cabeça. A formação dessas estruturas em regiões mais caudais é regulada pelo gene BRACHYURY (T). A lateralidade (assimetria esquerda-direita) é regulada por uma cascata de moléculas e de genes sinalizadores. FGF8, secretado por células no nó e na linha primitivos, induz a expressão de NODAL e de LEFTY2 do lado esquerdo, e esses genes aumentam a expressão de PITX2, um fator de transcrição e gene-chave para a lateralidade esquerda.

O neurotransmissor serotonina (5-HT) também atua na definição da lateralidade por meio da restrição da expressão de NODAL ao lado esquerdo. O posicionamento normal dos órgãos (à esquerda e à direita) é chamado de situs solitus, enquanto sua inversão completa é chamada de situs inversus. Quando um ou mais órgãos estiverem em posição anormal, essa condição será chamada de situs ambiguous ou heterotaxia. Indivíduos com situs inversus correm baixo risco de apresentar outros defeitos congênitos, mas seus filhos correm risco mais elevado, principalmente para cardiopatia. Em contrapartida, os pacientes com heterotaxia correm risco elevado de ter muitos tipos de malformações congênitas e quase todos apresentam algum tipo de anormalidade cardíaca. Erros nos níveis de 5-HT ou a expressão incorreta de genes da via de sinalização da lateralidade, como PITX2, resultam em defeitos de lateralidade, como dextrocardia, situs inversus e anomalias cardíacas. As mutações no fator de transcrição dedo de zinco ZIC3 causam heterotaxia associada ao X com cardiopatias graves e outros defeitos.
As células epiblásticas que se movem pelo nó e pela linha primitivos são predeterminadas por suas posições a se tornarem tipos específicos de mesoderma e de endoderma. Assim, é possível elaborar um mapa de destino do epiblasto mostrando seu padrão.
No fim da terceira semana, são estabelecidas três camadas germinativas na região da cabeça, consistindo em ectoderma, mesoderma e endoderma, e esse processo continua a produzir essas camadas para as áreas mais caudais do embrião até o fim da quarta semana. A diferenciação tecidual e orgânica já começou e ocorre no sentido cefalocaudal à medida que a gastrulação continua.
Enquanto isso, o trofoblasto progride rapidamente. As vilosidades primárias adquirem um centro mesenquimatoso, no qual surgem pequenos capilares. Quando esses capilares vilosos fazem contato com os capilares da placa coriônica e do pedúnculo embrionário, o sistema viloso está pronto para fornecer nutrientes e oxigênio ao embrião.
Anotações aula
Corpo lúteo é responsável pela produção da progesterona, para desenvolver endométrio e produção de muco cervical.
Muco cervical produzido pela progesterona facilita a entrada do espermatozoide, favorecendo o ambiente para fecundação. O muco facilita a propulsão do espermatozoide pela melhora na viscosidade, evitando que ele retroaja.
Progesterona faz o feedback negativo em cima da hipófise, diminuindo sua produção. Altos índices sustentados de progesterona faz com que gere uma espécie de tampão onde o espermatozoide não é favorecido a penetração.
Progesterona baixa não faz efeito nenhum, progesterona em nível médio faz efeito positivo para fecundidade, se o nível estiver muito alto gera feedback negativo, logo anticoncepção.
FSH é responsável pelo desenvolvimento e diferenciação do folículo. LH faz oocitacao e age na maturação do corpo lúteo. Crescimento do endométrio é favorecido pela progesterona.
Gonadotrofina coriônica é o hormônio que mantém o corpo lúteo ativo, até o quarto mês, quando é substituído pela placenta.
Estrogênio é produzido principalmente pelas células da granulosa (granulamentosas). A partir da oocitação tem o rompimento das células da granulosa fazendo uma diferenciação que estimula a progesterona, fazendo com que o corpo prepare o endométrio para uma possível fertilização.
Quando o corpo lúteo se degenera há menstruação.
Se a mulher engravidar a progesterona não cai, se mantém em um patamar alto para manter esse embrião.
Quando o espermatozoide é produzido, ele é produzido nos testículos em uma região chamada nos túbulos seminíferos, se acumulando no epidídimo.
Substâncias são secretadas no líquido seminal (proteínas) para ajudar a resistir o PH vaginal. Algumas substâncias contidas na cabeça do espermatozoide ajudando na fecundação.
A zona pelúcida é formada por glicoproteínas, já acima da zona pelúcida existe um resquício das células da granulosa do folículo chamada corona radiada.
A capacitação, período no qual uma capa de glicoproteínas e de proteínas plasmáticas seminais é removida da cabeça do espermatozoide, faz com que algumas proteínas do líquido seminal, que estão temporariamente bloqueando essa região, se desprendam, possibilitando a ligação entre o acrossomo e a zona pelúcida. Essa ligação favorece a liberação de substâncias semelhantes à acrosina e à tripsina, que vão ajudar com que as glicoproteínas da zona pelúcida sejam quebradas, tornando o oócito mais permeável e a partir das enzimas lisossomais dos grânulos corticais, que são hidrolíticas, quebram a membrana possibilitando a entrada do espermatozoide no oócito.
Durante a fertilização, o espermatozoide precisa penetrar na:
Zona pelúcida
Camada de glicoproteínas que cerca o oócito, facilita e mantém a ligação do espermatozoide e induz a reação acrossômica. A permeabilidade da zona pelúcida se altera quando a cabeça do espermatozoide contata a superfície do oócito (despolarização que torna a zona pelúcida rígida), liberando enzimas lisossomais dos grânulos corticais que por sua vez alteram as propriedades da zona pelúcida, evitando a possibilidade de penetração de qualquer outro esperatozoide.
Após os pró-núcleos se fundirem (cariogamia) formando o zigoto (fusão dos pro-nucleos femininos e masculinos formando o zigoto 46 cromossomos), eles replicam seus DNAs, onde os cromossomos paterno e materno se misturam, dividindo-se longitudinalmente e passando por uma divisão mitótica, que dá origem ao estágio de duas células.
Os resultados da fertilização são a restauração do número diploide dos cromossomos, a determinação do sexo cromossômico e o início da clivagem.
Ampola da tuba uterina, local mais provável para acontecer a fertilização, é onde o zigoto é gerado. Esse zigoto que estava na ampola da tuba uterina vai continuar o deslocamento até chegar ao útero.
Gêmeos monozigóticos, se originam a partir de um zigoto que se reparte ao meio gerando duas clivagens. Material genético idêntico.
Gêmeos dizigóticos, dois ovócitos diferentes são fecundados (por exemplo em casos de liberação de ovócitos pelas duas tubas uterinas concomitantemente) com material genético diverso, gerando dois corpos completamente diferentes em material genético.
Zona pelúcida limita a expansão na clivagem, dividindo as células exponencialmente, porém essas diminuindo de tamanho. Quando elas formam uma massa celular, recebe o nome de mórula (entre 8 e 16 células).
A mórula começa a sair da tuba uterina e entrar no útero no 6 dia de gestação, encontrando um ambiente adequado para a implantação.
Por volta do período que a mórula entra na cavidade uterina, ela sofre diferenciação e forma uma estrutura embrionária chamado blastocisto. O blastocisto surge quando começa a penetrar líquido uterino (líquido rico em glicogênio) onde essa mórula absorve esse liquido, separando dois grupos celulares em embrioblasto (massa interna) e trofoblasto (massa externa), e a cavidade blastocística. Blastocisto se caracteriza pela divisão com duas massas e uma cavidade interna.
Enzimas do trofoblasto começa a digerir para realizar a nidação. Trofoblasto para de existir para da origem ao sinciciotrofoblasto e citrotrofoblasto.
É nessa fase que há implantação no endométrio (nidação).
Embrioblasto dará origem ao embrião, já o trofoblasto irá formar a placenta. Para que esse embrião em formação adentre no estroma uterino precisa de um endométrio preparado, espesso, com glândulas ativas, vasos disponíveis para nutrir o embrião, ou seja o endométrio precisa estar em aspecto esponjoso que é encontrado na fase secretora.
Na formação do embrião as células do trofoblasto vão começar a ganhar algumas diferenças, sendo que uma parte vai penetrar rumo ao estroma uterino, e outra parte vai definir relações referente a placenta.
Se diferencia em citotrofoblasto (vai permanecer dando limitação ao blastocisto e fazendo diferenciação dando origem em novos tecidos) e sinciciotrofoblasto (penetrar o epitélio endometrial dando condução a implantação e adentrando cada vez mais no estroma).
Segunda semana de gestação
O blastocisto já está totalmente alojado no endométrio, já penetrou totalmente e o embrioblasto (não o trofoblasto que se transformou em sinciciotrofoblasto e citrotrofoblasto que gera a placenta e anexos) já está diferenciando nos seguintes folhetos embrionários:
-Epiblasto e o Hipoblasto
Exoceloma – camada células da parte interna que vem do hipoblasto e que forma saco vitelínico que no futuro se torna saco coriônico que irá formar a placenta. Com a evolução o saco vitelínico tende a ser pequeno, enquanto em aves é gigante, pois a parte nutricional vai ser dado pela placenta.
Olhar para o número de folhetos embrionários para saber em qual momento da gravidez está.
Blastocisto – 2 folhetos
Gastrula – 3 folhetos
Gastrula já começa a formação dos eixos.
A gatrulação é o processo que estabelece as 3 camadas (folhetos) germinativas (ectoderma, mesoderma e endoderma) no embrião. Esse evento tem inicio com a formação da linha primitiva
Epiblasto passa a ser ectoderma
Mesoderma também do epiblasto
Hipoblasto passa a ser endoderma
Fase lútea é a fase secretora para manter progesterona no útero, e o que mantém a fase secretora é a progesterona.
O corpo lúteo vai produzir beta hcg para manter essa progesterona, para que assim não haja aborto. Posteriormente quem vai continuar produzindo a progesterona será a placenta.
Linhagem evolutiva na fecundação –> Zigoto –> Blastomeros –> Mórula –> Blastocisto –> Gastrula
No final da aula fomos ao laboratório de histologia para ver laminas. Fiz esse esquema com todas as fases principais:

REFLEXÃO: Essa aula foi muito boa, didática e produtiva. Neander nos explicou sobre coisas que acontecem na ovulação concomitantemente, e que não haviamos entendido. Elucidou muitas duvidas e se colocou a disposição para nos ajudar em explicações caso a gente se perca nos conceitos. Isso foi bastante importante pois nos trouxe não só clareza as informações mas também maior segurança para nós, aquela "luz no fim do tunel". Estamos bem esperçosos que agora as coisas "engrenem" mais e mais.



Comentários